Problem 7: Konvergent evolusjon ødelegger logikken bak felles avstamning
Oversatt herfra.
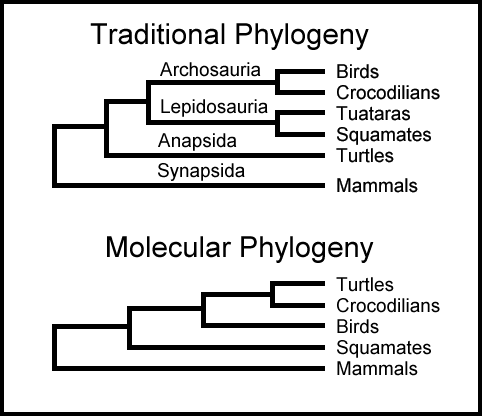 I problem 6 så vi at hoved-forutsetningen bak alle fylogenetiske trær er at biologisk likhet er et resultat av arv fra en felles stamfar. Problemet for evolusjonære biologer som står overfor motstridende evolusjonære trær, er at biologisk likhet ofte opptrer på steder som ikke er forutsatt av vanlig avstamning. Med andre ord anerkjenner alle at biologiske likheter ofte forekommer blant arter, i tilfeller der de ikke kan forklares som et resultat av arv fra en felles stamfar. Dette betyr at hoved-forutsetningen feiler.
I problem 6 så vi at hoved-forutsetningen bak alle fylogenetiske trær er at biologisk likhet er et resultat av arv fra en felles stamfar. Problemet for evolusjonære biologer som står overfor motstridende evolusjonære trær, er at biologisk likhet ofte opptrer på steder som ikke er forutsatt av vanlig avstamning. Med andre ord anerkjenner alle at biologiske likheter ofte forekommer blant arter, i tilfeller der de ikke kan forklares som et resultat av arv fra en felles stamfar. Dette betyr at hoved-forutsetningen feiler.
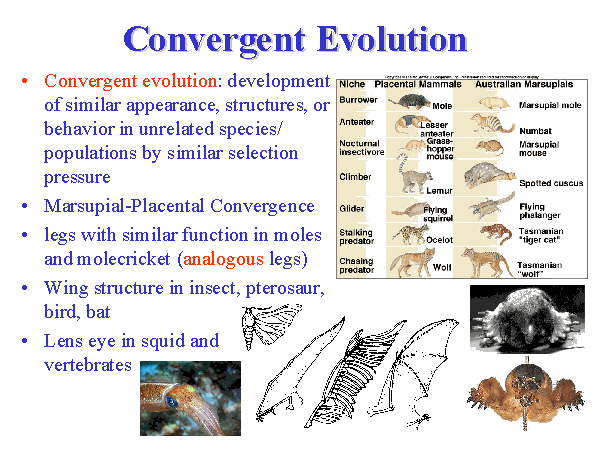
Vi så også på slutten av Problem 6 at når biologer ikke klarer å konstruere fylogenetiske trær, gjør de ofte ad hoc appeller til andre prosesser, for å forklare data som ikke passer til et trelike mønstre. En av disse forklaringene er konvergent evolusjon, hvor evolusjonære biologer postulerer at organismer oppnår de samme egenskapene selvstendig, i separate linjer, og ikke gjennom arv fra en felles stamfar. Når evolusjonære biologer blir tvunget til å appellere til konvergent evolusjon, reflekterer det et sammenbrudd i hovedforutsetningen, og en manglende evne i å tilpasse dataene til et trelikt mønster. Eksempler på dette florerer i litteraturen, men noen få vil være tilstrekkelig.
Genetisk konvergent evolusjon
En artikkel i Journal of Molecular Evolution fant at molekylbaserte fylogenier var i konflikt med tidligere etablerte fylogenier av store pattedyrgrupper, og konkluderte med at dette uregelmessige treet "ikke skyldes en stokastisk feil, men skyldes konvergent eller parallell evolusjon." (119)
En studie i Proceedings fra U.S. National Academy of Sciences forklarer at når biologer forsøkte å konstruere et fylogenetisk tre for de største gruppene av fugler ved hjelp av mitokondrialt DNA (mtDNA), var deres resultater kraftig i konflikt med tradisjonelle forestillinger om fuglerelasjoner. De fant til og med 'konvergent' likhet mellom noen fugle-mtDNA og mtDNA av fjerne arter som slanger og øgler. Artikkelen antyder at fugle-mtDNA gjennomgikk 'flere uavhengige opprinnelser', med studien som foreslår "flere uavhengige opprinnelser til en bestemt mtDNA-gen sammenheng blant diverse fugler." (120)
Et 2005-artikkel i Nature Immunology bemerket at planter og dyr har en svært lik biokjemisk organisering av deres respektive medfødte immunsystem, men deres felles stamfar hadde ikke et slikt immunsystem:
-Selv om det ser ut til å være generelt akseptert at de medfødte immunresponsene til planter og dyr deler i det minste noen felles evolusjonære opprinnelser, feiler undersøkelse av tilgjengelige data å støtte denne konklusjonen, til tross for likheter i den overordnede logikken til den medfødte immunresponsen i mangfoldige multi-cellulære [organismer]. (121)
Ifølge artikkelen kan ikke felles avstamning forklare disse 'uventet lignende' systemene 'som tyder på uavhengig evolusjonær opprinnelse i planter og dyr.' Artikkelen er tvunget til å konkludere med at slike komplekse likheter gir et 'overbevisende tilfelle av 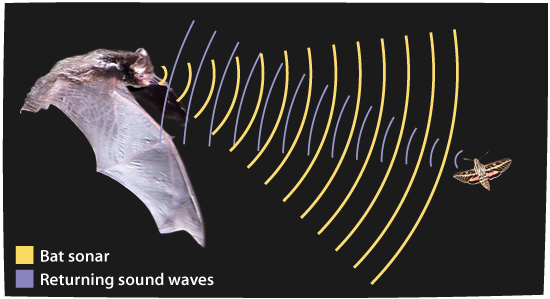 konvergent utvikling av medfødte immunforsvar.' (122)
konvergent utvikling av medfødte immunforsvar.' (122)
Et annet kjent eksempel på konvergent evolusjon er evnen til flaggermus og hval til å bruke ekkolokalisering, selv om deres fjerne, vanlige forfedre ikke hadde denne egenskapen. Evolusjonære biologer trodde lang tid at dette var et tilfelle av morfologisk konvergens, men en artikkel i Current Biology forklarer det 'overraskende' funnet at ekkolokalisering i flaggermus og hval også innebærer genetisk konvergens:
-Bare mikro flaggermus og tannhvaler har oppnådd sofistikert ekkolokalisering, uunnværlig for orientering og matfangst. Selv om flaggermus- og hval -bio-sonarene oppsto uavhengig og avviker vesentlig i mange aspekter, rapporterer vi det overraskende funnet at delfin, en tannhval, er gruppert med mikro-flaggermus i gentreet, konstruert ved hjelp av proteinsekvenser kodet av høre-genet prestin. (123)
En artikkel kalt disse dataene, 'ett av de beste eksemplene på konvergerende molekylær evolusjon oppdaget til dags dato.' (124) Men igjen er disse neppe isolerte eksempler. I 2010 forklarte en artikkel i Trends in Genetics:
 -Den nylige brede bruken av genetiske og/eller fylogenetiske tilnærminger har avdekket mangfoldige eksempler på gjentatt evolusjon av adaptive trekk, inkludert flere fremtoninger av øyne, ekkolokalisering i flaggermus og delfiner, pigment-modifikasjoner hos virveldyr, etterligning i sommerfugler for gjensidig interaksjon, konvergens av noen blomster-egenskaper i planter, og flere uavhengige utviklinger av bestemte proteinegenskaper. (125)
-Den nylige brede bruken av genetiske og/eller fylogenetiske tilnærminger har avdekket mangfoldige eksempler på gjentatt evolusjon av adaptive trekk, inkludert flere fremtoninger av øyne, ekkolokalisering i flaggermus og delfiner, pigment-modifikasjoner hos virveldyr, etterligning i sommerfugler for gjensidig interaksjon, konvergens av noen blomster-egenskaper i planter, og flere uavhengige utviklinger av bestemte proteinegenskaper. (125)
Biokjemiker og Darwin-skeptiker Fazale Rana gjennomgikk den tekniske litteraturen og dokumenterte over 100 rapporterte tilfeller av konvergent genetisk utvikling. (126) Hvert tilfelle viser et eksempel hvor biologisk likhet - selv på genetisk nivå - ikke er et resultat av arv fra en felles stamfar. Så hva gjør dette med hoved-forutsetningen for trebygging at biologisk likhet innebærer arv fra en felles stamfar?
Jorden er rund, men er felles avstamning troverdig?
 En evolusjonær forsker prøvde å presse leserne til å akseptere darwinismen ved å hevde at "biologer i dag vurderer at felles avstamning for hele livet, er et faktum på linje med jordens kuleform." (127) Men er slike kategoriske uttalelser engang nyttige, og i enda mindre grad sanne?
En evolusjonær forsker prøvde å presse leserne til å akseptere darwinismen ved å hevde at "biologer i dag vurderer at felles avstamning for hele livet, er et faktum på linje med jordens kuleform." (127) Men er slike kategoriske uttalelser engang nyttige, og i enda mindre grad sanne?
Talsmenn for neo-darwinistisk evolusjon blir tvunget til å argumentere for at biologisk likhet innebærer felles avstamning, bortsett fra når den ikke gjør det. Og i de mange tilfeller der den ikke gjør det, appellerer de til alle slags ad-hoc-rasjonaliseringer for å redde felles avstamning.
Talende nok er den ene antakelsen det sjelden settes spørsmålstegn ved, selve den generelle antakelsen om felles avstamning. Men kanskje årsaken til at forskjellige gener forteller forskjellige evolusjonære historier, er at gener har helt forskjellige historier å fortelle, nemlig historier som indikerer at alle organismer ikke er genetisk relaterte. Det er noe håp om en annen historie som er mer tilpasset dataene, som Michael Syvanen våget å foreslå i Annual Review of Genetics 2012, at "livet kan faktisk ha flere opprinnelser." (128) Med andre ord kan universell felles avstamning faktisk, ikke være sant.
Referanser:
 [119]. Ying Cao, Axel Janke, Peter J. Waddell, Michael Westerman, Osamu Takenaka, Shigenori Murata, Norihiro Okada, Svante Pääbo, Masami Hasegawa, "Conflict Among Individual Mitochondrial Proteins in Resolving the Phylogeny of Eutherian Orders," Journal of Molecular Evolution, 47 (1998): 307-322.
[119]. Ying Cao, Axel Janke, Peter J. Waddell, Michael Westerman, Osamu Takenaka, Shigenori Murata, Norihiro Okada, Svante Pääbo, Masami Hasegawa, "Conflict Among Individual Mitochondrial Proteins in Resolving the Phylogeny of Eutherian Orders," Journal of Molecular Evolution, 47 (1998): 307-322.
[120.] David P. Mindell, Michael D. Sorenson, and Derek E. Dimcheff, "Multiple independent origins of mitochondrial gene order in birds," Proceedings of the National Academy of Sciences USA, 95 (September, 1998): 10693-10697.
[121.] Frederick M Ausubel, "Are innate immune signaling pathways in plants and animals conserved?," Nature Immunology, 6 (10): 973-979 (October, 2005). [122.] Ibid.
[123.] Ying Li, Zhen Liu, Peng Shi, and Jianzhi Zhang, "The hearing gene Prestin unites echolocating bats and whales," Current Biology, 20(2):R55-R56 (January, 2010) (internal citations removed).
[124.] Gareth Jones, "Molecular Evolution: Gene Convergence in Echolocating Mammals," Current Biology, 20(2):R62-R64 (January, 2010); Yong-Yi Shen, Lu Liang, Gui-Sheng Li, Robert W. Murphy, Ya-Ping Zhang, "Parallel Evolution of Auditory Genes for Echolocation in Bats and Toothed Whales," PLoS Genetics, 8 (6): e1002788 (June, 2012).
[125.] Pascal-Antoine Christin, Daniel M. Weinreich, and Guillaume Besnard, "Causes and evolutionary significance of genetic convergence," Trends in Genetics, 26(9):400-405 (2010) (internal citations omitted).
[126.] See Fazale Rana, The Cell’s Design: How Chemistry Reveals the Creator’s Artistry, pp. 207-214 (Baker Books, 2008).
[127.] Karl W. Giberson, Saving Darwin: How to be a Christian and Believe in Evolution, p. 53 (HarperOne, 2008).
[128.] Michael Syvanen, "Evolutionary Implications of Horizontal Gene Transfer," Annual Review of Genetics, 46:339-356 (2012).
Oversettelse og bilder ved Asbjørn E. Lund